Please put an active hyperlink to our site (www.rusnature.info) when you copy the materials from this page
Biomes and Regions of Northern Eurasia
Arid Environments
<<< Geological Structures and Major
Characteristics of Relief | Biomes & Regions Index
| Human Impact on Ecosystems >>>
Biota and Soils
Semi-desert Vegetation
The semi-desert biome is as an ecotone between the dry steppe and desert zones (Walter
and Box, 1983). Both vegetation types occur here side-by-side under the same macroclimatic
regime, and the dominance of one or the other vegetation type results from microclimatic
and litho-edaphic conditions. Increasingly arid from north to south and from west to east,
the regional climate favours the development of semi-desert biomes on the Caspian lowland
from the Terek valley to the Volga valley between 46°N and 50°N, then between 48°N and
50°N on the Volga-Ural watershed, and further east in Kazakhstan, approximately between
48°N and 49°N.
Despite the similar climatic conditions of the semi-deserts on the Caspian lowland and
Kazakh Knolls, their vegetation composition is very different and exhibits a strong
association with landforms and substrates. On the flat Caspian lowland, with its numerous
shallow endoreic lakes and shallow ground water lenses, the vegetation and soil patterns
typically follow the microrelief, which is a combination of microdepressions, and
micro-summits, and microslopes between them. Steppe, semi-desert or desert communities
develop here side-by-side following the microclimatic conditions.
Steppe vegetation is usually associated with micro-depressions, which occupy up to 25
per cent of the area. They accumulate more snow in winter, and in spring the soil receives
about 200 mm of snow melt water. The meadow chernozem-like soils here are usually
salt-free. The vegetation cover is typically dominated by Festuca sulcata, Stipa
lessingiana, S. capillata, Koeleria cristata, and Agropyron pectiniforme. The total
vegetation cover reaches 75-90 per cent and the average height of the plants is about
27-37 cm (Walter and Box, 1983). On the microslopes, which occupy about 25 per cent of the
Caspian semi-desert, one finds slightly alkaline, chestnut earths, with a plant cover
composed primarily of Tanacetwn achilleifolium and Agropyron desertorum with considerable
participation of Poa bulbosa. About 50-60 per cent of the Caspian semi-deserts is occupied
by the drier, desert-like communities dominated by Artemisia pauciflora and Kochia
prostrata or Camphorosma mon-speliacum. The total vegetation cover ranges from 20 per cent
to 45 per cent, and the height of plants is about 15-20 cm.
Such a three-member mosaic is just the general pattern of semi-desert vegetation in
this area. Four- and five-member mosaic structures also occur, when the vegetation cover
comprises solonetz soils, which are formed in shallow depressions because of rapid water
evaporation. Artemisia pauciflora, Camphorosma mon-speliacum, and Anabasis salsa occur on
crusted solonetz soils, while Kochia prostrata and Artemisia lercheana are typical of deep
columnar solonetz soils. Formation of such a micromosaic is quite typical of young, very
level, slightly saline maritime sediments.
The halobiomes occur around salt lakes, where the ground water can rise to the surface
by capillarity and evaporate. The most extreme solonchak sites, as for example around Lake
Baskunchak, consist of hard sodium chloride and carry no vegetation. The first pioneer
species are generally Salicornia europaea and Halocnemum strobilaceum.
In central Kazakhstan, the typical vegetation of the semi-desert zone is composed of
two associations dominated by Stipa, Festuca sulcata, and Artemisia gracilescens. The
flora of the Kazakh deserts and semi-deserts is genetically related to the Mongolian and
Turanian deserts. The richness of the flora decreases with increasing dryness. The number
of species identified for the semi-desert of Kazakhstan is 423 (Walter and Box, 1983), and
the number of Holarctic species decreases from north to south, while the number of
Mediterranean species, which are typical of the Iran-Turanian deserts, increases in the
same direction.
The zonal soil type in the semi-desert is a light chestnut earth, which represents the
transition between the true chestnut earths of the dry steppe and the brown earths of the
desert. A typical feature of the semi-desert vegetation in Kazakhstan is domination of
Artemisia gracilescens, a typically Kazakh flora element. Festuca sulcata continuously
decreases southward and is completely lacking at the southern boundary of the semi-desert
(Walter and Box, 1983).
There are considerable variations of semi-desert vegetation in different litho-edaphic
conditions. Psammophytic vegetation is found on the sandy river terraces of eastern
Kazakhstan. It mainly consists of Stipetum pennatae and Festucetum beckerii, the latter on
mobile sands. Most widespread are hemi-psammophytic variants of the typical vegetation,
which contain Artemisia marshalliana, psammophytes Gypsophila paniculata and Syrenia
ses-siliflora, and plants adapted to both stony and sandy soils, Ephedra distachya and
Hulthemia berberidiflora, as well as some steppe species. Petrophytes appear on shallow
and rocky soils. The most common species are Stipa sareptana and Artemisia gracilescens,
mostly in complexes with the solonetz communities (Atriplex ńŕďŕ, Anabasis salsa, and
Nanophyton spp.). Artemisia frigida together with Allium galanthum, Anabasis truncata, and
Sedum albertii are typical on rubble surfaces. Rocky soils are usually covered with shrubs
on the metamorphic rocks of northern Kazakhstan. Halophytes are very widespread in the
semi-desert zone. A particularly characteristic species is Atriplex ccma, which occurs in
association with Artemisia species. Stands of the very tall grass Lasiagrostis splendens
are especially typical on slightly saline soils with a high ground water level. On
non-saline, moist to wet sites, plant communities include herbaceous plants similar to
those in the steppe. Sites with calcareous light chestnut earths or calcareous solonetz
soils are dominated by Anabasis salsa, Suaeda physophora, and Atraphaxis decipiens.
Characteristic plants for calcareous soils are Linosyris tatarica and species of
Serratula.
Desert Vegetation
Desert biome occupies the entire area between latitudes 38°N and 44°N and longitudes
55°E and 65°E. This corresponds latitudinally to the northern Mediterranean zone. Desert
conditions also occur on the Caspian lowland and in the Kazakh Knolls south from 48-49°N,
as well as on the Apsheron peninsula in Azerbaijan. There are, however, significant
floristic differences between these regions, and especially between northern and southern
desert biomes.
The so-called northern deserts occur on the Caspian lowland, in central and eastern
Kazakhstan, and in Transcaucasia. The Caspian sandy desert is a subbiome which lies below
sea level (Yakubov, 1955). Although brown earths are the zonal soil type, most of this
area is covered by sands. The sands are underlain by heavy loams, and the ground water
table is mostly quite high. On the sands the plant cover is dominated by Artemisia
lercheana, Agropyron spp., Festuca sulcata, and Koekria glauca. Tamarix is found in the
depressions between sand hills and on the Caspian coast Kalidium caspicum and Halostachys
caspica occur. Groups of Salix appear in the reed stands on the coast.
The desert vegetation of central and eastern Kazakhstan is distinguished by the absence
of grasses and by the presence of more xerophilous Artemisia species and various desert
dwarf shrubs. Salsola arbusculiformis is a very important species, occuring with Artemisia
but also with Ferula ferulaeoides and Caragana balchaschensis. Ephemerals are quite
numerous. In the more petrophytic variants of the Salsola community, some dwarf shrubs,
forbs, and grasses also appear (Walter and Box, 1983).
The desert zone of the Kazakh Knolls continues to the south as the Betpak-Dala desert.
The most important dominant plant species here are Salsola laridfolia, Artemisia
terra-albae, and Anabasis salsa. Ephemerals are almost completely absent from the Kazakh
deserts. This is due to both the rocky or crusted soils and a climate with cold winters
and dry springs. Halophytic communities in the salinized depressions are similar to those
of the Caspian lowland and include species of Anabasis, Atriplex ńŕďŕ, Artemisia
pauciflora, and Halocnemum strobilaceum (Walter and Box, 1983).
The flora of the Turanian deserts is composed primarily of the Iran-Turanian species,
which also tend towards the Mediterranean (Korovin, 1962; Walter and Box, 1983). In
contrast to Kazakhstan, Mongolian species are absent in this zone. The level of endemism
is very high, as the result of intensive mountain formation in the late Tertiary,
increased aridity, and isolation from neighbouring regions (Rustamov, 1994).
The southern deserts of Central Asia feature stronger aridity and milder winters. As a
result, the humus content in soils is very low and is only found in the uppermost horizon.
The high evaporation rate produces a crust on the soil surface. These soils are rich in
calcium and especially in gypsum, which explains the wide expansion of gypsum-tolerant
vegetation in the deserts of the Turanian lowland. The following genera are represented in
Central Asia by numerous species: Calligonum (67), Ammodendron (26), Zygophyllum (31),
Anabasis (20), Salsola (54), Limonium (22), Tamarix (16), and Suaeda (22). Gypsum-tolerant
dwarf-shrub vegetation on grey soils of plateau sites includes many endemic taxa and also
species in the genera Cleome, Reaumuria, Haplophyllum, Ferula, FJorema, Arthrophytum, and
Nanophyton, as well as semi-shrubs such as Artemisia turanica.
Psammophytic communities of Central Asia include 350 species, 56 per cent of which are
endemics. The genus Calligonum with thirty species has its centre of distribution in
Mongolia. These are joined by five Ammodendron species, the twenty closely related
Astragalus species, Haloxylon persicum, three Salsola species, the endemic Smirnovia
turkestana, and Ephedra. The virgate form is characteristic of the plants of sandy deserts
with good water supplies. Therophytes are quite numerous: a total of 143 species, of which
98 are spring ephemerals. Of the ephemeroids, thirty-one species are present, including
Carex physodes, Poa bulbosa, eight species of Allium, two species of Tulipa, four species
of Ferula, and many others (Plate 12.1).

Plate 12.1 Vegetation of the Karakum desert
Halophytic desert vegetation is represented in Central Asia by many genera with a great
species diversity: Salsola (20), Suaeda (9), Atriplex (4), Pandorea (2), Camphorosma (3),
Echinopsilon (3), Kochia (2), Kalidium (3), Halostachys, Halocnemum, Salicornia,
Seidlitzia, Anabasis (8), Haloxylon, Petrosimonia (10), Halocharis (3), Halimo-cnemis (2),
Halogeton (2), Zygophyllum (14), Nitraria (3), Limonium (9), and Reaumuria (3). This type
of vegetation is restricted to solonchak soils close to ground water and thus accompanies
the lower courses of rivers, occupies depressions, and encircles saline lakes. Most
species are succulents, with high osmotic values caused by the chlorides stored in the
cell fluids.
The takyr environments, which occur in the Amudarya and Syrdarya deltas and on the
piedmonts of the Kopetdagh mountains, are often thought to be totally free of vegetation
but they are densely populated by algae and lichens (Rodin, 1977). The takyrs are flooded
sometimes several times during the year, however, the water quickly evaporates from the
surface leaving the dried-out clay. While higher plants are either completely absent or
remain relatively unimportant, a total of 147 algae species were identified on the takyr
sites of Central Asia with algae FJesertialgeta and lichens Desertilichineta being
dominant species (Rodin, 1961: Walter and Box, 1983). The lichen communities develop
alongside the algae communities but on the somewhat higher non-flooded sites because, in
contrast to algae, lichens need a dry environment where there is no sedimentation. The
takyr surfaces are subject to continuous changes resulting from flooding and
sedimentation. The vegetation of inundated areas is continuously changing and the algae
and lichen communities are in a dynamic state, replacing each other whenever the
distribution of the water changes. Microscopic soil fungi, such as Dematiaceae, are also
widespread on the takyr sites (Litvinov, 1956).
Floodplain Vegetation
All the larger permanent streams, which rise in the mountains and carry water to the
deserts, are accompanied by varied and very dynamic floodplain vegetation. According to
Rodin (1963), these include floodplain forests (locally termed tugai), shrub thickets,
tall grass and forb vegetation (termed jangil), meadows, and the areas of the most
recently deposited soil (termed kair).
The most important tugai tree species are Populus diversifolia and P. pruinosa. These
trees are relatively short-lived reaching an age of 35-45 years and a height of 13-15 m.
Underneath the tree tier one can find understory trees and shrubs 3-7 m high, which
include Elaeagnus orientalis and species of Salix. The true shrub tier is 2-4 m high and
contains Tamarix species and Hcdimodendron holodendron. Vines are richly developed. The
herb layer has fewer species and its composition varies with soil wetness, salt content of
the soil, and the availability of light on the forest floor. The tugai forests typically
occupy areas, which are inundated annually for a short time, and where the ground water
level stays at a depth of 4-7 m.
Another type of floodplain vegetation is shrub thickets which are composed of several
Tamarix species. Most commonly shrubs are 5-6 m tall. In contrast to the. floodplain
forests, the shrub thickets are not inundated and occur on sites with ground water at a
depth of 4-5 m. On increasingly dry sites, Tamarix thickets form a transition to the
desert-like tree vegetation with Haloxylon ammodendron (Figure 12.5).
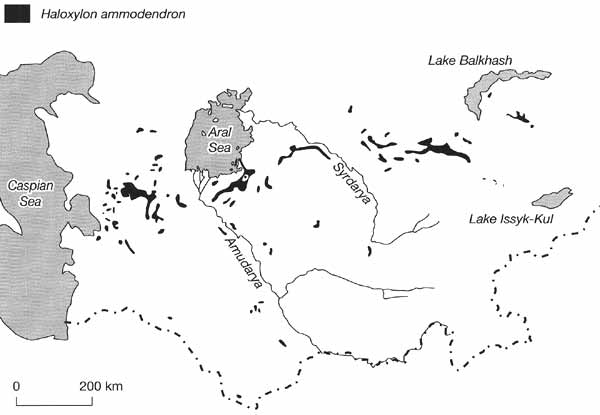
Fig. 12.5 Areas of the extensive distribution of Haloxylon ammodendron.
Modified from Walter and Box (1983)
In the delta region of the Amudarya and in the shallow water of the Aral Sea,
communities with Phragmites spp. form the so-called jangil, impenetrable reed tall-grass
thickets. Floodplain meadows occupy large areas in the south-west and in the delta areas
of the Amudarya and the Atrek, but also occur in the foothills of the Kopetdagh. The most
common here are Aeluropus stands and ephemerals. Finally, the areas of recently deposited
soil, with very little vegetation, consisting of pioneer plants (Typha pallida and
Calamagrostis dubia), also occur on the floodplains.
Fauna
Most of the faunal data for deserts and semi-deserts of the FSU are of the
zoogeographic type, with general data for the particular species, their distribution and
frequency, etc. The significance of the individual species in food chains, which is
complicated, is hardly mentioned. Quantitative data, which are seldom available for
well-studied ecosystems, are lacking for Kazakhstan and most of Central Asia.
The fauna of Central Asia and Kazakhstan has been modified by human impact, especially
grazing, which has been continuing here for centuries. Today, the wild horses (tarpans),
which were common here a century ago, are extinct. The population of the Asiatic wild ass
(kulan) was reduced dramatically and only massive efforts at preservation and restoration
of this species in nature reserves has prevented its extinction. The population of the
wild antelopes (saiga), which is found in the wild in a few areas of Kazakhstan, has also
declined. In order to protect the herds, the carnivores were exterminated, destroying the
natural equilibrium in the process. The rodents profited from this action and are today
the most numerous mammals — carnivorous birds also benefited.
The most common species in the desert and semi-desert ecosystems are rodents: among the
29 mammal species that occur in central Kazakhstan, 22 are rodents. For most species the
semi-desert is a transitional zone: 20 mammal species (14 rodents) are desert species and
8 (6 rodents) are steppe species. Other semi-desert species include carnivores (e.g.,
weasel), insectivores, and bats. In the northern deserts of Kazakhstan, rodents dominate
but in the protected areas one can find ungulates (e.g., saiga and the Persian gazelle),
carnivores (e.g., polecat, fox, and spotted wildcat), and insectivores. Of the insects of
Kazakhstan several groups have been studied in detail: Homoptera: Euchenorryncha
(plant-sucking cicada, represented here by 365 species) and Psylloidea (leaf lice, 91
species); Coleoptera represented by 357 beetle species; Hymenoptera: Braconidae (ichneumon
flies, 268 species); Diptera: Chloropidae (75 species); and Lepidoptera (butterflies, 338
species) (Walter and Box, 1983).
Among the southern deserts of Central Asia the most detailed data on fauna exist for
the Karakum desert. As in the northern deserts, rodents are the largest group and are of
great ecological importance because of their impact on plants (Stalmakova, 1954; Rodin,
1977). Their diet consists of fruits, seeds, and to a lesser extent herbaceous parts of
plants. As a result of the precarious water balance, the rodents of the southern deserts
reproduce only during the short favourable period in March-April. If these species occur
in the wetter semi-desert zone, they generally have two or three reproductive cycles per
year. In the protected areas the desert hare is widespread. Among the carnivores, the
mouse-weasel and polecat are common. The canines are represented by the fox, as well as by
jackal near oases along the Murgab and Amudarya rivers, and rarely by wolf. Of the cats,
the spotted wildcat is very common. The ungulates are represented by wild pigs who migrate
from the floodplain forests to sandy desert when the ephemerals develop in spring, and in
protected areas, by the Persian gazelle.
Among other vertebrates one should mention bird species, which are found in all
communities (e.g., raven, golden eagle, little owl, and long-legged buzzard). The reptile
species are equally common in the mobile sands, for example, the sand boa (Eryx miliaris),
species of racerunner (Eremias spp.), and in the Haloxylon stands, for example, agama
(Agama sanguinolenta), racerunner (Eremias carinata), racer (Coluber turia), and tortoise
(Testudo horsfieldii). Geckos (Gymnodactylus russwi and G. caspicus), arrow-snake
(Psammophis dueviatus), and desert monitor (Varanus griseus) are common across the
Karakum. Most of the reptiles are carnivorous, especially snakes, which greatly reduces
the rodent population.
Among the invertebrates of the desert zone, those that are closely related to the woody
plants, are much better studied. The Haloxylon ammodendron stands are a habitat for many
groups of invertebrates, in the majority phytophagous. A very common carnivorous group are
spiders (Araneae), which represent in summer up to 3 6 per cent of the total zoomass of
all invertebrates in the canopy (Walter and Box, 1983). The invertebrate soil fauna
include carnivorous scorpions, pseudo-scorpions, and spiders. Phytophagous and
detritovorous groups include Trombiformes, Thysanoptera, the larvae and imagines of
beetles, and the larvae of Diptera. Among animals which spend most of their time
underground, the most noteworthy are the bright-coloured ants of the genus Camponotus and
the soil cicadas of the genus Pentasterdius. The large termites, whose mounds reach 50-100
cm in height, are restricted to the Kopetdagh piedmonts.
<<< Geological Structures and Major
Characteristics of Relief | Biomes & Regions Index
| Human Impact on Ecosystems >>>
Contents of the Arid Environments
section:
Other sections of Biomes & Regions:
|
|







