Please put an active hyperlink to our site (www.rusnature.info) when you copy the materials from this page
Physical Geography of Northern Eurasia
Biodiversity and Productivity of Ecosystems
<<< Biomes of Northern Eurasia and
Climate | Physical Geography Index | Geography
of Diversity of Animal Populations >>>
Geography of Diversity of Plant Communities
Figure 7.6 illustrates the taxonomic diversity of flora in Northern Eurasia.
Undisturbed protected areas (nature reserves) were used for the assessment of richness of
vascular plant species.
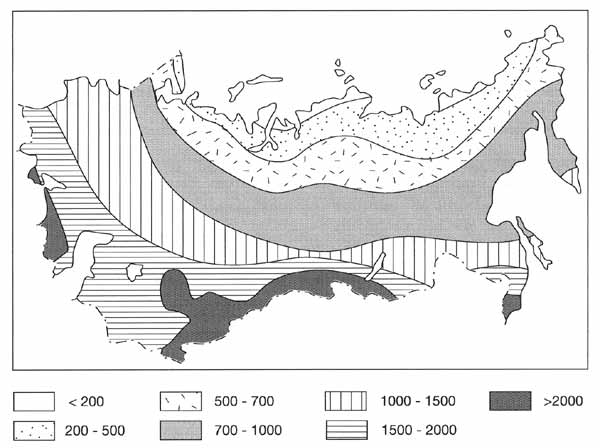
Fig. 7.6 Number of species of vascular plants on a standard area of 100
km2. After Malyshev (1992)
The number of species, computed for a standard area of 100 square kilometres, varies
greatly from less than 200 in polar deserts to more than 3000 species in the semi-arid and
arid landscapes (including steppes, semi-deserts, and deserts) of southern European
Russia, Transcaucasia, and Central Asia.
In the polar deserts, the young age of landscapes and the extremely low temperatures
are responsible for the formation of skeletal soils and low biodiversity. Vegetation
communities are dominated by lower plants (mainly algae and lichens) with the inclusion of
relatively few flowering species such as Papaver polare and species of Saxifraga,
Puccinellia, Poa, and Luzula. The number of vascular plants in local floras usually varies
between twenty at the northern edge of polar deserts and sixty in the south (Aleksandrova,
1988).
The tundra biome covers extensive areas, reaching its greatest latitudinal extent in
the circumpolar Arctic on the Taymyr peninsula, where all polar biomes (from the polar
deserts to tundra-taiga ecotone) are present. An important feature of tundra is the
absence of trees in zonal habitats. Mosses dominate among plants and play a vital role in
the protection of permafrost. Flowering plants, grasses, and sedges are more widespread
than in the polar deserts and dwarf shrubs (Betula nana, Betula exillis, Salix spp.) occur
across typical and southern tundra. In southern tundra, bush tier develops on watersheds.
Boreal elements of flora are prominent in the vegetation communities of southern tundra
and that makes it similar to the forest-tundra ecotone. Tundra is a relatively young biome
and therefore, the species richness and endemism are generally low. The highest level of
endemism occurs in Eastern Siberia and the Chukchi peninsula which have not been fully
glaciated (Aleksandrova, 1980). Elements of the ancient Pleistocene flora, at present
found mainly in the steppes of Southern Siberia and Mongolia, survived in the
tundra-steppe associations of the Chuckchi peninsula (Yurtsev, 1981). However, the major
components of the current Chukotkan flora are plants, requiring wet and poorly drained
soils (Eriophorum vaginatum, Carex lugens, Ledum palustre), which were probably uncommon
in the Pleistocene. Vegetation of the Chukotkan tundra, therefore, is a complex mosaic of
the tundra-steppe, tussock, and moss-lichen communities. Another region with relatively
high species richness and endemism is Wrangel island, which was at different times
isolated from and connected to the continent (Yurtsev, 1994; Chernov and Matveeva, 1997).
Boreal coniferous forests (taiga) are remarkable for their vast extent. However,
despite the high diversity of landscapes, the European and Siberian taiga is very
species-poor in comparison, for example, with the coniferous forests of North America
(Walter, 1980). Only a few tree species dominate the taiga: Picea abies, Picea obovata,
Abies sibirica, Pinus sibirica, Pinus sylvestris, and Larix sibirica. Eastwards of the
Yenisey river, in Central and Eastern Siberia and on the Sea of Okhotsk coast, Larix
gmelinii becomes a dominant. It has a shallow root system, allowing it to grow on a thin
soil layer above the permafrost. The flora of the taiga lacks endemics and there are very
few rare plants. Larch forests are characterized by the lowest species-richness among
forest ecosystems with their flora comprising not more than 450 species of vascular plants
(see below).
The biome of mixed broad-leaved coniferous forests does not form a continuous zone but
occurs on the East European plain and in the Far East. There are different views on
attributing mixed and deciduous forest ecosystems to biomes. Traditionally, mixed and
deciduous forests have been treated as two separate zones (Lavrenko and Sochava, 1956).
Later it was suggested that only one biome of mixed forests should be recognized (Milkov
and Gvozdetsky, 1976) and deciduous forests, together with meadow steppes, should be
viewed as a part of the forest-steppe zone (Milkov, 1986) (this issue is addressed below).
The main tree species in mixed and deciduous forests belong to the taxa of Tilia, Quercus,
Fagus, Carpinus, Fraxinus, Acer, Pinus, and Picea. In the north and west of the mixed
forest biome, conifers dominate while in the south deciduous species are more important.
Mixed forests of the East European plain are characterized by a higher species diversity
than the taiga. However, in comparison with the mixed forests of eastern Asia, the
floristic composition of the European forests is poor mainly due to the depletion of
species by repeated glaciations. In contrast, the mixed forests of the Far East are much
more species-rich. A less extreme environmental history has produced a complex flora which
includes both Siberian and Manchurian species. There are many rare species and about 200
endemics (Alpatiev et al., 1973).
Although the word 'steppe' creates an image of meagre and uniform vegetation, in fact
the opposite is true of the European steppes and forest-steppes. These are the most
fertile areas of Northern Eurasia and the biological diversity of these biomes is very
high. Forest-steppe is a mosaic of woodlands and meadow steppes. In the European
territory, forests are formed mainly by deciduous broad-leaved taxa (mainly Quercus and
Tilia) while in Western Siberia Betula dominates. Eastwards of the Altay mountains, Larix
is the most common tree species and here the forest-steppe forms an ecotone between the
coniferous boreal forests and the open steppes. Local floras of the European forest-steppe
grasslands contain over a thousand species of vascular plants per 100 km2
(Tishkov, 1995) or up to 40-50 species per m2 (Dokhman, 1968; Lavrenko and Karamysheva,
1993). Common species include Bromus, Flomis, Filipendula, Galium, Poa, Stipa, Salvia, and
Trifolium. Southwards and eastwards, in more arid conditions, species richness declines to
12-15 species per m2 (Dokhman, 1968; Lavrenko and Karamysheva, 1993). Here, in the true
and dry steppes, species of Festuca, Helictotrichon, Koeleria, Poa, and Stipa dominate and
species of Artemisia become common (Titlyanova et al., 1990).
Forest-steppe and steppes, especially in the East European plain, have been exposed to
enormous stress and disturbance mainly through the development of agriculture. Reviewing
grasslands of Europe, Knapp (1979) commented that natural steppes originally extended
across the southern European FSU and to the dry plains of Romania, Bulgaria, Hungary, and
former Yugoslavia, while at present only small fragments of natural ecosystems survive.
Arid ecosystems of semi-desert and desert biomes occupy about 11 per cent of the
territory of the FSU (Isachenko, 1985) and occur on the Caspian lowland, in Transcaucasia,
Kazakhstan, and Central Asia. [In this chapter, the term 'Central Asia' refers to the
republics of Central Asia (Turkmenistan, Tajikistan, Uzbekistan, and Kyrgyzstan). In the
Russian and German-language literature, deserts of this region are often termed Middle
Asian, in contrast to the Central Asian, which refer to the deserts of Mongolia, China,
and Afghanistan].
Apart from Transcaucasia and the south of the Turanian lowland, the arid biomes of
Northern Eurasia belong to the temperate climate zone and are distinguished from the
subtropical deserts and semi-deserts by cold winters with often very low temperatures
produced by the Siberian high.
Vegetation of semi-deserts has a complex character due to the transitional position of
this biome between steppes and deserts. However, species-richness is much lower than in
steppes and there are about 150-200 species of vascular plants per 100 km2
(Bazilevich et al., 1986), mainly species of Artemisia, grasses (Agropyron, Botriochloa,
Festuca, and Stipa) and annuals. Salt-tolerant vegetation develops around saline lakes,
such as the Elton and the Baskunchak on the Caspian lowland, where the ground water table
is so high that water can rise to the surface by capillarity and evaporate (Walter and
Box, 1983a). In contrast, the semi-deserts of Transcaucasia are almost Mediterranean in
character. Along gradients of increasing rainfall, they used to develop into open
woodlands with single-standing trees of Pistacia mutica and Juniperus spp. These
woodlands, however, have been almost totally wiped out (Walter and Box, 1983a).
In deserts, there are about 100-150 species per 100 km2 (Bazilevich et al.,
1986). The zonal vegetation is distinguished from that of semi-deserts by the absence of
grasses and by the domination of the more xerophilous Artemisia species. There are many
classifications of deserts and desert ecosystems. The following main types are singled
out: sandy, ephemeral, gypsum, halophyte deserts, and takyrs, the bare clayey lands which
are flooded in spring and dry out and crack into polygon sections in summer (Walter and
Box, 1983b). In the conditions of high continentality and aridity, types of substrate and
ground water availability control the character and distribution of vegetation and,
because of it, vegetation types correspond to soil types (i.e., gypsum-rich, takyr). Sandy
deserts, the Karakum and the Kyzylkum, occupy vast areas on the Turanian lowland between
the Caspian Sea and the Syrdarya. Woody plants (e.g., Haloxylon spp. and Acacia spp.) and
low shrubs (Artemisia, Salsola, and Anabasis) dominate their flora. As a result of their
isolation from the neighboring regions by the Sarmat Sea and the intensive formation of
the mountains, which created a biogeographical barrier in the late Tertiary, this area is
distinguished by well-developed endemism (Walter and Box, 1983fc). In gypsum deserts,
vegetation consists mainly of gypsum-tolerant plants. There are about 400 gypsum-tolerant
species on the Turanian lowland (Korovin, 1961). The halophyte deserts develop where water
is close to the surface, although the decisive ecological factor here is not water but
rather salts, primarily chlorides. Few plants are adapted to the high salt concentrations
and the most common species are hydrohalophytes: Salicornia herbacea, Halocnemum
strobilaceum, Haloxylon ammodendron, and endemic Seidlitzia rosmarinus. The ephemeral
deserts develop on loessic, salt-free soils. The vegetation period lasts from March to May
and during this time annual species develop, the most common of which are Carex hostiana
and Đîŕ bulbosa. In those springs when rainfall is relatively high, ephemeral deserts
have the appearance of a meadow. The number of species in the ephemeral desert is 45-50
(Walter and Box, 1983b). Widespread in the entire desert zone are takyrs. For a long time,
takyrs were thought to be totally without vegetation although it is now known that they
are colonized by algae and lichens. The algae may be of a very old, perhaps pre-Devonian
type (Walter and Box, 1983b). Higher plants, which do occur in takyrs, are annuals which
germinate from seeds brought by wind and spring water and take hold in hollows formed by
water erosion and filled with soil which is also carried by spring water and wind.
There are four centres of species diversity, all of which are located in mountainous
regions: the Crimea and Caucasus, Central Asia (mountains of the Tien-Shan and Pamir-Alay
systems), Altay and Sayans in Southern Siberia, and the Maritime Province (the
Sikhote-Alin) in the Pacific sector.
Two major factors are responsible for the high level of plant species richness in the
mountains: historical (mountains served as refugia where species were able to survive the
unfavorable climatic epochs such as glaciations and sea transgressions) and ecological
(i.e., greater variety of habitats). The species-richness of the Caucasus is remarkably
high. There are more than 6000 plant species concentrated in a geographically small area,
whereas in the whole of the European part of Northern Eurasia there are only about 3500
species (Milkov and Gvozdetsky, 1976). The unique nature of the Caucasian flora is
discussed in the regional description part.
Plant communities always include a number of endemics and relics (i.e., species rare in
themselves). However, many species become rare and endangered if their habitats are
disturbed, or they themselves become overexploited (e.g., by overcutting of forests or
overgrazing of grasslands). A register of endangered species, the so-called Red Data Book
(1984), contains 533 species of plants in need of protection of which 440 species (or 82
per cent) are angiosperms, 11 species are gymnosperms, 11 are ferns, 4 are club mosses, 22
are mosses, 29 are lichens, and 17 are fungi. The number of rare and endangered species is
particularly high in the southern regions where it exceeds 20 per cent of all species.
Using Red Data Book information, Martynov and Volkov (1994) compiled a map illustrating
the distribution of rare and endangered species (Figure 7.7).
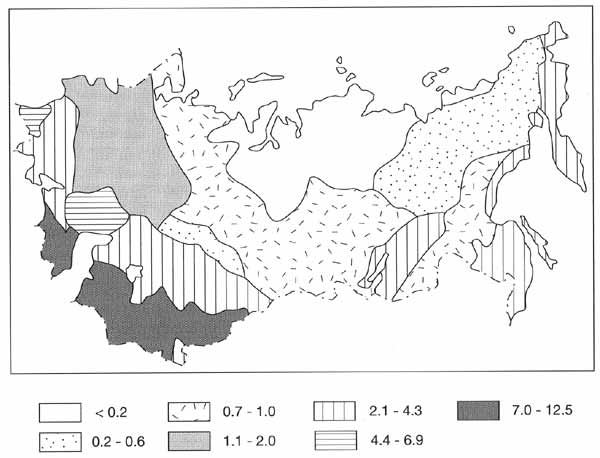
Fig. 7.7 Rare plant species included in the Red Data Book as percentage
of the total number of plant species. After Martynov an Volkov (1994)
The number of endangered species correlates with the total number of species of
vascular plants in regional floras (Figure 7.6).
<<< Biomes of Northern Eurasia and
Climate | Physical Geography Index | Geography
of Diversity of Animal Populations >>>
|







